 对照。第7天观察是否长菌落。
对照。第7天观察是否长菌落。
1.2.2 对峙培养 供试菌株扩大培养及打取菌饼方法同“1.2.1”节中的方法,采用对峙培养法[7],将木霉与病菌两两组合,同时接种于无菌PDA平板(直径90 mm)中,2个接种点相距 45 mm,以不接种木霉菌株的PDA培养基作对照,设3次重复,在28 ℃条件下恒温培养,第7天测定病菌的菌落直径,计算其生长抑制率。
抑制率=[(dCK-dB)/dCK]×100%。
式中:dCK表示对照病菌菌落直径;dB表示处理病菌菌落直径。
1.2.3 数据统计 所有试验数据均采用SPSS 19.0统计软件Duncans多重比较法进行统计分析,计算处理间的差异显著性。
2 结果与分析
2.1 原始木霉与抗药性木霉菌株菌落特征
木霉菌J1、J3、Y2菌株经含扑海因杀菌剂PDA平板培养,获得4株抗药性木霉菌菌株,分别为PJ1-1、PJ1-2(J1的抗药性菌株)、PJ3(J3的抗药性菌株)、PY2(Y2的抗药性菌株),抗药性菌株比原始菌株的菌落颜色浅(图1)。
2.2 各供试木霉菌菌株对枇杷根腐病病菌P3.1菌株的抑制作用
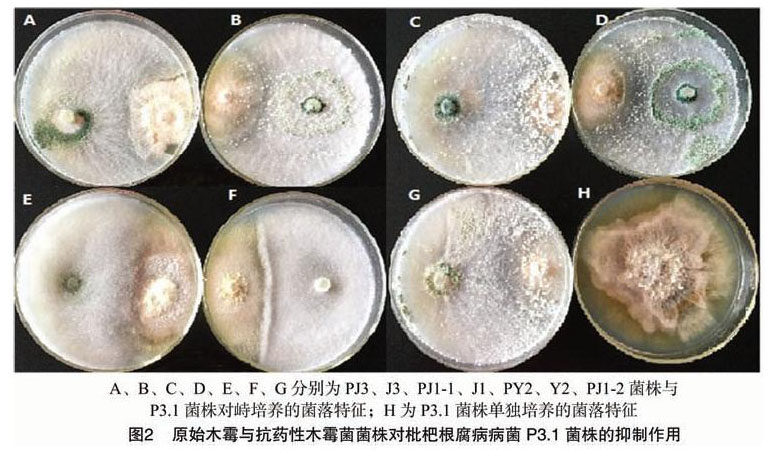
由表1可知,7个木霉菌株对枇杷根腐病病菌P3.1菌株的抑制率差异极显著(P<0.01)。抗药性菌株PJ1-1、PJ1-2、PJ3对枇杷根腐病病菌P3.1菌株的抑制率均高于57%,抑菌作用强于其原始菌株。抗药性菌株PY2对枇杷根腐病病菌P3.1菌株的抑制率略低于其原始菌株。说明抗药性菌株PJ1-1、PJ1-2、PJ3对枇杷根腐病病菌P3.1菌株的抑菌活性未衰退。其中,PJ1-1和PJ1-2抗药性菌株完全覆蓋枇杷根腐病病菌P3.1菌株,其上长出大量分生孢子,重寄生作用明显强于原始菌株(图2)。
2.3 各供试木霉菌菌株对枇杷根腐病病菌P3.5菌株的抑制作用
由表2可知,7个木霉菌菌株对枇杷根腐病病菌P3.5菌株的抑制率差异极显著(P<0.01)。抗药性菌株PJ1-1、PJ1-2、PJ3对枇杷根腐病病菌P3.5菌株的抑制率均高于50%,抑菌作用强于其原始菌株。抗药性菌株PY2对枇杷根腐病病菌P3.5菌株的抑制率极显著高于其原始菌株(P<0.01)。说明抗药性菌株 PJ1-1、PJ1-2、PJ3、PY2对枇杷根腐病病菌P3.5菌株的抑菌活性未衰退。其中,PJ1-1抗药性菌株的菌落完全覆盖枇杷根腐病病菌P3.5菌株,其上长出大量分生孢子,重寄生作用明显强于原始菌株(图3)。
2.4 各供试木霉菌菌株对枇杷根腐病病菌P3.6菌株的抑制作用
由表3可知,7个木霉菌菌株对枇杷根腐病病菌P3.6菌株的抑制率差异极显著(P<0.01)。抗药性菌株PJ1-1、PJ1-2、PJ3对枇杷根腐病病菌P3.6菌株的抑制率均高于60%,抑菌作用强于其原始菌株。抗药性菌株PY2对枇杷根腐病病菌P3.6菌株的抑制率低于其原始菌株,但产孢能力增强(图4)。说明抗药性菌株PJ1-1、PJ1-2、PJ3对枇杷根腐病病菌P3.6菌株的抑制活性未衰退。其中,抗药性菌株PJ1-1完全覆盖枇杷根腐病病菌P3.6菌株,其上长出大量分生孢子,重寄生作用明显强于其原始菌株(图4)。
2.5 各供试木霉菌菌株对石榴枯萎病病菌的抑菌作用
由表4可知,7个木霉菌菌株对石榴枯萎病病菌SK菌株的抑制率差异极显著(P<0.01)。抗药性菌株PJ1-1、PJ1-2、PJ3的抑制率均高于71%,抑菌作用强于其原始菌株。抗药性菌株PY2菌株对石榴枯萎病病菌SK菌株的抑制率极显著高于其原始菌株(P<0.01)。说明抗药性菌株 PJ1-1、PJ1-2、PJ3、PY2对石榴枯萎病病菌SK菌株的抑制活性未衰退。其中,抗药性菌株PJ1-1、PJ1-2、PJ3的产孢能力有所下降(图5)。
2.6 各供试木霉菌菌株对万寿菊叶斑病病菌的抑菌作用
由表5可知,7个木霉菌菌株对万寿菊叶斑病病菌WJ1菌株均具有不同程度的抑制作用。抗药性菌株PJ1-1、PJ1-2、PJ3对万寿菊叶斑病病菌WJ1菌株的抑制率均高于62%,抑菌作用强于其原始菌株。抗药性菌株PY2对万寿菊叶斑病病菌WJ1菌株的抑制率极显著低于其原始菌株。说明抗药性菌株PJ1-1、PJ1-2、PJ3对万寿菊叶斑病病菌WJ1菌株的抑制活性未衰退。其中,PJ1-1和PJ1-2抗药性菌株完全覆盖万寿菊叶斑病病菌WJ1菌株,其上长出大量分生孢子,重寄生作用明显强于其原始菌株(图6)。
3 结论与讨论
抗药性木霉菌株PJ1-1、PJ1-2、PJ3对供试的5株病原菌抑菌活性均高于原始菌株J1和J3,其中,PJ1-1菌株完全覆盖病原菌,其上产生大量分生孢子。说明其具有极强的重寄生能力,生防前景广阔。前人曾研究获得抗腐霉利[8]、抗苯菌灵[7]、多菌灵[9-10]、速克灵[11-12]、能降解敌敌畏[13]、毒死蜱和甲胺磷[14]等的抗药性木霉菌菌株。但筛选获得抗扑海因的木霉菌菌株尚属首次。
我国化肥和农药过量施用现象较严重,由此引起环境污染和农产品质量安全等重大问题。农作物绿色防控的呼声越来越高。木霉与化学农药混配使用能大大降低化学农药施用量,有时还能发挥协同增效作用。例如,段银芝等研究发现,吡虫啉、咪唑乙烟酸、咪唑烟酸等3种农药能促进哈茨木霉菌菌丝生长及产孢[15]。田连生等报道,木霉菌剂与多菌灵混配防治灰霉病有协同增效作用[16]。任凤山等发现,木霉与几种杀菌剂混配能增强对苹果轮纹病的防治效果[17]。木霉菌 PJ1-1 菌株与扑海因混配施用, 防治石榴枯萎病、枇杷根腐病和万寿菊叶斑病是否具有协同增效作用,有待进一步研究。
参考文献:
[1]刘云龙,何永宏,王新志. 国内一种果树新病害——石榴枯萎病[J]. 植物检疫,2003,17(4):206-208.
[2]Lu H J,Wang C M,Zheng X L,et al. First report of loquat root rot disease caused by Pestalotiopsis microspora in China[J]. Plant Disease,2016,100(5):1008.
[3]魯海菊,潘柳君,李 河,等. 云南万寿菊叶斑病病原菌的鉴定与ITS序列分析[J]. 西北农业学报,2015,24(6):116-119.
[4]王占斌,黄 哲,祝长龙. 木霉拮抗菌在植物病害生物防治上的应用[J]. 防护林科技,2007(4):104-107.
[5]杨合同,唐文华,Ryder M.木霉菌与植物病害的生物防治[J]. 山东科学,1999,12(4):7-15,20.
[6]于春生. 哈茨木霉菌多菌灵抗性选育与β-微管蛋白突变位点研究[D]. 大庆:黑龙江八一农垦大学,2015.
[7]周红姿,李宝聚,刘开启. 抗药性木霉菌研究进展[J]. 北方园艺,2003(6):10-11.
[8]丁 中,刘 峰,慕立义. 紫外光诱导哈茨木霉产生腐霉利抗性菌株的研究[J]. 中国生物防治,2002,18(2):75-78.
[9]杨 谦,赵小岩. 多菌灵抗性基因在木霉菌中的转化方法[J]. 科学通报,1998,43(22):2423-2426.
[10]鲁海菊,刘云龙,张云霞,等. 哈茨木霉多菌灵耐药性菌株的筛选[J]. 云南农业大学学报,2005,20(3):436-437.
[11]王 勇,杨秀荣,刘水芳. 拮抗木霉耐药性菌株的筛选及其与速克灵防治灰霉病的协同作用[J]. 天津农学院学报,2002,9(4):19-22.
[12]尹 婷. 深绿木霉T2对微生物种群数量的影响及抗药菌株的筛选[D]. 兰州:甘肃农业大学,2012.
[13]付文祥. 有机磷农药降解菌木霉FM10的生长条件研究[J]. 生物磁学,2005,5(3):29-31.
[14]刘 新,尤民生,魏英智,等. 木霉Y对毒死蜱和甲胺磷的降解作用[J]. 福建农林大学学报(自然科学版),2002,31(4):455-458.
[15]段银芝,郎剑锋,孔凡彬,等. 4种农药对哈茨木霉生长的影响[J]. 江苏农业科学,2013,41(3):101-102.
[16]田连生,陈 菲. 木霉菌剂与多菌灵协同防治灰霉病试验[J]. 江苏农业科学,2013,41(12):132-133.
[17]任凤山,王 燕,翟一凡,等. 木霉与几种化学杀菌剂协同防治苹果轮纹病[J]. 北方园艺,2015(16):111-115.